
By Adrian Wang Xinting, University of Oxford
Citation
Wang, A.W. (2021) ‘Natural laboratories’ in perspective: a review of literature on theories of island biogeography. Routes 2(1): 15-22
Abstract
Islands are diverse, well-defined, and relatively uncomplicated, facilitating significant progress in the study of ecological and evolutionary processes. MacArthur and Wilson’s (1963) Equilibrium Theory of Island Biogeography (ETIB) was a revolutionary framework that facilitated the understanding of the biogeographical processes in insular ecosystems. Over time, more sophisticated theoretical models have sought to advance the field of island biogeography, with Whittaker et al.’s (2008) General Dynamic Model (GDM) being a distinguished example, linking island age with species richness. However, there are several factors that may influence the accuracy and reliability of island studies, such as palaeo-configurations, anthropogenic activity, and evolutionary syndromes. This literature review serves as a synthesis of critiques from published scientific journals on the use of models in insular biogeography.
Glossary
Endemism: The state of a species being native to a single defined geographic location.
Insular biota: A term used to refer to animal, plant, and other organism life relating to, dwelling, or situated on an island.
Phenotypic divergence: A term which describes variation within the population in terms of the observable characteristics of an organism that result from the interaction of its genotype with the environment.
Adaptive radiation: The process in which organisms undergo rapid diversification from a single ancestral species into a range of new species that exhibit ecological traits specialised to different niches.
Species-area relationship: A term that describes the general pattern of increase in species richness with an increasing area of a habitat, or of part of the area of observation.
1. Introduction
Islands are the ‘logical laboratories of biogeography and evolution’, with each insular habitat being ‘an experiment awaiting the analysis of evolution and ecology’.
Edward O. Wilson, cited in Losos et al., 2010, p.7.
Diverse, well-defined, and relatively uncomplicated, islands are mediums through which one can study the diversification of species as they adapt to different landscapes (Kohler 2002a, cited in Hennessy, 2018). Moreover, as they vary in features such as area and degree of isolation, islands serve as scientific experimental systems that allow for the study of evolutionary responses of community properties (Lomolino et al., 2006). This essay will explore the utility of islands as natural laboratories for the study of ecological, evolutionary, and biogeographical processes, through models that predict and explain endemism.
Serving as a literature review, it will systematically examine the temporal development of island biogeographical models and their prominent features and characteristics. Simultaneously, it surfaces potential weaknesses of these models that may influence the accuracy and reliability of these findings, such as not taking into account palaeo-configurations, anthropogenic activity, and evolutionary syndromes.
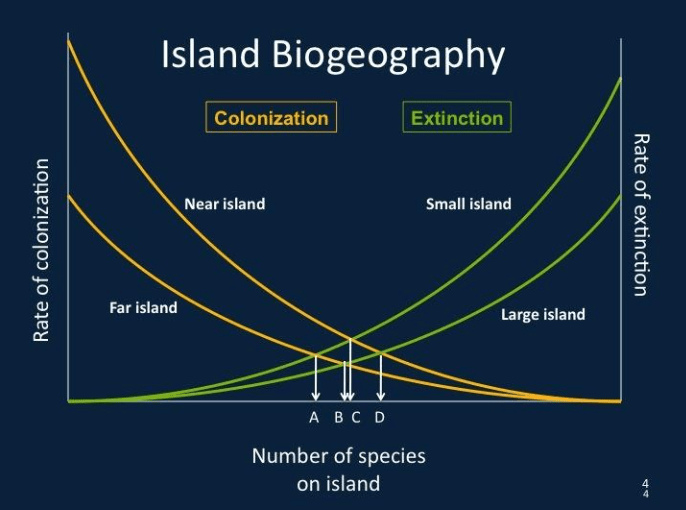
2. Equilibrium Theory of Island Biogeography
The Equilibrium Theory of Island Biogeography (ETIB) proposed by MacArthur & Wilson (1963) posits that the species richness of an island represents a dynamic equilibrium between opposing rates of immigration and extinction (see Figure 1). The ETIB is grounded on three fundamental premises of insular biotas: a positive correlation in the species-area relationship, species number decreasing with increasing isolation, and there being a continual turnover of species over time (Lomolino et al., 2010). Consequently, it is supposedly able to predict the relative rates at which islands of ‘different sizes and degrees of isolation should return to equilibrium, in the event that the biota is perturbed’ (ibid, p. 524). For instance, a near-source island would return to equilibrium sooner as compared to a more distant island of the same size, due to a disparity in immigration rate, assuming a constant extinction mechanism for both islands (ibid). Furthermore, the ETIB hypothesises that the size and relative isolation of the island affects local endemism. Larger islands, with lower extinction rates, permit the accumulation of endemics, while more isolated islands, through the ‘radiation zone’ effect, encourage greater proportions of endemism (Borregaard et al., 2016).
However, critics of the ETIB highlight how it does not account for several crucial factors that determine the evolutionary processes of diversification and extinction within remote islands (Borregaard et al., 2016). Firstly, it is argued that ETIB utilises island area as a sweeping indicator of islands’ ability to sustain its species richness. As a result, the model disregards the disparities in habitat prerequisites for different species of organisms, which are crucial in the carrying capacity of similar islands (Lomolino et al., 2010). This is evident in the presence of ‘specialist’ species, whose habitat specifications may only be fulfilled on sizeable islands could further facilitate the diversification in richness (ibid). Moreover, the origins of an island biota may be difficult to determine without examining the ‘systematics and historical distribution of the species that are present’, with existing species on an insular biota resulting from a range of factors, such as ‘over-water dispersal from continents and other islands, historical links to other islands and in situ speciation’ (ibid, p. 527). Finally, the ETIB omits the presence of disturbances, which contributes to setting the population off equilibrium. In the occurrence of major environmental disturbances such as volcanic eruptions and hurricanes, insular populations may never achieve equilibrium, or momentarily at most (Heaney, 2000, cited in ibid).
3. General Dynamic Model
A comparatively more integrated theory based on oceanic island biogeography was consequently found in the General Dynamic Model (GDM) proposed by Whittaker et al. (2008). By incorporating the precepts of the ETIB within an evolutionary time-scale, it highlights the increasingly influential roles of within-island speciation and geological dynamics (Whittaker et al., 2008). The GDM posits that the physical geographical attributes of oceanic islands demonstrate predictable dynamics over evolutionary time-scales, inducing developments in the evolutionary and ecological adaptations of species in island biotas (Borregaard et al., 2016). As the islands age, volcanic activity diminishes and they lose elevation by erosion and subsidence. This leads to the local extinction of species adapted to high-elevation habitats and lower nutrient availability in surfaces and soil (Price & Clague, 2002). Effectively, the GDM argues that the biotas of islands are determined by the relative rates of immigration, local extinction and speciation, in accordance with the phases of emergence, development, and submergence of island development (Whittaker et al., 2008).
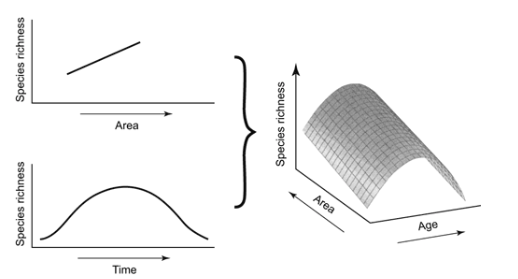
a unimodal relationship with time and a linear rise with increasing area.
In contrast to the ETIB, the GDM suggests that islands’ distinct, dynamic geology have a considerable impact on their habitat complexity and speciation over time (Borregaard et al., 2017). Biodiversity indices, such as species richness and endemism, are influenced through developments in the area, elevation, and environment of the island as it ages. This is manifested in a unimodal/ hump-shaped trend in the carrying capacity of the island over its life cycle (see Figure 2). Moreover, Borregaard et al. (2016) posits that the evolutionary rates are augmented through two mechanisms – the area, elevation and topographic complexity of the island determine the overall size of the resource base, while the topographical complexity of the island influences the likelihood of reproductive isolation of species populations within the island (p. 808). In terms of diversity-dependent dynamics, the GDM asserts that an environmental carrying capacity restricts the species richness that can be supported on an island at each stage (Borregaard et al., 2017). The initial periods of the island’s ontogeny entails the resource base surpassing the realized species richness, due to there being insufficient time for in situ speciation to mature. This untapped ecological potential stimulates adaptive radiation, promoting productivity before species richness reaches a plateau (ibid). Eventually, the resource base declines and the island founders, as speciation rates decline and extinction rates climb (Whittaker et al., 2008).
In addition, Borregaard et al. (2016) found that low immigration rates on remote islands meant that realised richness will invariably remain below resource supply levels for much of the life cycle of the island. This is evidenced by the phenotypic divergence present on remote islands – an instance of a diversity-dependent response to unoccupied niche space – as mainland species adapted to particular ecological opportunities have yet to immigrate (Losos & Ricklefs, 2009, cited in Borregaard et al., 2016). Consequently, it implies that over much of an island’s lifespan, the system is arguably not in a dynamic equilibrium, contrary to the assumption implied by the ETIB.
4. Not necessarily an ideal laboratory
Yet, the results from experiments conducted on islands may not be widely applicable nor transferable. These ‘natural laboratories’ influence their taxa via evolutionary syndromes, as insular environments induce ‘fundamental transformations in the character of insular species’ (Lomolino, 2010: 986). The loss of dispersal prowess, a propensity towards flightlessness, and developing naïveté toward predators are examples of converging traits of insularization (Whittaker et al., 2017). For instance, flightlessness in birds is manifested through an increased emphasis on forelimbs, as a ‘direct response to reduced predation pressure in the absence of terrestrial mammals and specialist bird-hunting avian predators’ (ibid, p. 5). Consequently, these reversals in natural selection left many insular species especially vulnerable to introduced vertebrate predators, causing the former to be threatened with extinction.
Moreover, islands may not be apt natural laboratories due to anthropogenic influence. Through the means of transport, urbanization, and agriculture, human presence has significantly modified the structure and ecology of local species in the forms of insular colonization, habitat fragmentation, and dispersal of invasive species (van de Geer et al., 2017). For example, the introduction of commensal mammals, such as pigs (Sus domesticus) and goats (Capra hircus), has had a major negative effect on vegetation and has been associated with the extinction of native species in Macaronesia (Nogué et al., 2017). These anthropogenic disturbances have been immense, with Steadman (2006) estimating that of the initial 500 to 1600 species of flightless rails that once inhabited the Pacific Islands, only 16 remain currently. Furthermore, Nogué et al. (2017) posits that human colonization and the resulting cultural activity may have even influenced evolutionary processes through genetic divergence, causing the evolution of unique island species and subspecies. The Canarian Egyptian vulture (Neophron percnopterus) was found to have colonised the Canary Islands under anthropogenic facilitation through the ‘introduction of domesticated livestock, as suitable food sources were previously lacking’ (Agudo et al., 2010, cited in Nogué et al., 2017). As a result, van de Geer et al. (2017) has found that ‘longstanding models of island biogeography may prove inadequate unless their conceptual domains are expanded to include … the impact of human activities on each of these processes’ (p. 1003).
Alternatively, instead of island models, palaeo-configurations may be a better method for the study of biogeographical processes. Norder et al. (2018) found that extreme archipelago configurations that have prevailed during the Last Glacial Maximum (LGM) are ‘not sufficient to explain endemism patterns on volcanic oceanic islands’ (p. 192). Instead, palaeo-configurations from the Pleistocene, observable at intermediate sea levels, left a superior imprint on single-island endemic richness sequences. Interglacial ice-melt and sea-level rise – induced by climatic fluctuations – caused the depletion of ecosystems at higher elevations, impacting extinction, speciation and biota size (ibid). Thus, a more extensive temporal scale may be more apt in understanding communities’ relationships.
5. Conclusions
In conclusion, islands are model ecosystems for biogeographic processes, such as trophic relationships and species dynamics. Inferences from scientific studies could further understandings of human-environment interactions, especially in the field of biodiversity conservation, amidst increasing anthropogenic impact. However, as MacDonald et al. (2018) posits, there is a substantial range of factors that ‘operate simultaneously to structure species diversity in insular and fragmented systems’ (p. 2741). Given the significance of evolutionary syndromes, human influence, and the recency of island configurations on insular speciation, islands may not necessarily serve as models for optimal natural laboratories. Moreover, Hennessy (2018) argues that designating archipelagos as ‘natural laboratories’ may be controversial, as it intrinsically encompasses the ‘conservationist territorial and biopolitical management of nature’ (p. 500). Future research on islands could seek to further understand the evolutionary dynamics of insular biotas across a range of dissimilar archipelagos under contemporary developments, such as climate change and pollution. It could also seek to develop enhanced models that take into account anomalies or extremities. An instance of this is the Small Island Effect, which occurs when island area falls below a certain threshold. It involves an extensive disruption of the species-area relationship, with species being determined by habitat instead (Triantis et al., 2006).
| Theory | Key Contributions | Critiques |
| ETIB (MacArthur & Wilson, 1963) | Species richness of an island varies with area and isolation – a dynamic equilibrium between opposing rates of immigration and extinction. Species richness increases with an increasing area of a habitat. Greater habitat heterogeneity induces reduced extinction rates. Simultaneously, richness decreases with rising island isolation, due to diminishing immigration rates. | Establishes key foundations of island biogeography. However, overly simplistic, omitting factors such as differences in habitat conditions and the manifestation of disturbances. |
| GDM (Whittaker et al., 2008) | Positive relationship between species richness and geological age of island, together with area. Species richness increases linearly with island area, while demonstrating a a unimodal relationship with time. Environmental variables influence level of endemism in islands. | Widely applicable. Given the theory’s focus on oceanic islands, a potential area of future research could involve specific niches in insular habitats. |
6. Acknowledgement
I am extremely thankful for the thorough feedback and support of Dr Marc Macias-Fauria and the two anonymous reviewers of this essay.
7. References
Borregaard, M.K., Matthews, T.J. & Whittaker, R.J. (2016) The general dynamic model: towards a unified theory of island biogeography? Global Ecology and Biogeography 25(7), 805-816. DOI: 10.1111/geb.12348
Borregaard, M.K., Amorim, I.R., Borges, P.A.V., Cabral, J.S., Fernández-Palacios, J.M., Field, R., Heaney, L.R., Kreft, H. , Matthews, T.J., Olesen, J.M., Price, J., Rigal, F., Diver, Kim C. (2008) Not as the crow flies: assessing effective isolation for island biogeographical analysis. Journal of biogeography, 35(6), pp.1040–1048.
Heaney, L.R. (2000) Dynamic disequilibrium: a long‐term, large‐scale perspective on the equilibrium model of island biogeography. Global Ecology and Biogeography, 9(1), pp.59–74.
Hennessy, Elizabeth, 2018. The politics of a natural laboratory: Claiming territory and governing life in the Galápagos Islands. Social Studies Of Science, 48(4), pp.pp483–506.
Lomolino, M.V., Riddle, B.R. & Brown, J.H. (2006) Biogeography, 3rd edn. Sinauer Associates, Sunderland, MA.
Lomolino M.V., Riddle B.R., Whittaker R.J. & Brown J.H. (2010) Biogeography (4th edition), Sinauer Associates, Inc., Sunderland – Chapters 13 & 14.
Lomolino M.V. (2010) Four Darwinian themes on the origin, evolution and preservation of island life. Journal of Biogeography 37, 985–994. DOI: 10.1111/j.1365-2699.2009.02247.x
Losos, J.B. & Ricklefs, R.E. (2009) Adaptation and diversification on islands. Nature, 457(7231), pp.830–836.
Losos, J.B., Ricklefs, R.E. & MacArthur, R.H. (2010) The theory of island biogeography revisited, Princeton: Princeton University Press.
MacArthur, R.H. & Wilson, E.O. (1963) The theory of island biogeography, Princeton: Princeton University Press.
MacDonald, Z.G., Anderson, I.D., Acorn, J.H. and Nielsen, S.E., 2018. The theory of island biogeography, the sample‐area effect, and the habitat diversity hypothesis: complementarity in a naturally fragmented landscape of lake islands. Journal of Biogeography, 45(12), pp.2730-2743.
Nogué, S., de Nascimento, L., Froyd, C. A., Wilmshurst, J. M., de Boer, E. J., Coffey, E. E. D., Whittaker, R. J., Fernández-Palacios, J. M., Willis, K. J. (2017) Island biodiversity conservation needs palaeoecology. Nature Ecology & Evolution 1(7), 181. DOI: 10.1038/s41559-017-0181
Norder, S. J., Proios, K., Whittaker, R. J., Alonso, M. R., Borges, P. A. V., Borregaard, M. K., . . . Rijsdijk, K. F. (2018) Beyond the Last Glacial Maximum: Island endemism is best explained by long-lasting archipelago configurations. Global Ecology and Biogeography 28(2), 184-197. DOI:10.1111/geb.12835
Price, J.P. & Clague, D.A. (2002) How old is the Hawaiian biota Geology and phylogeny suggest recent divergence. Proceedings of the Royal Society B: Biological Sciences, 269(1508), pp.2429–2435.
Steadman, D.W. (2006) Extinction & biogeography of tropical Pacific birds, Chicago ; London: University of Chicago Press.
Steinbauer, M.J., Triantis, K.A. ,Valente, L. ,Weigelt, P. & Whittaker, R.J. (2017) Oceanic island biogeography through the lens of the general dynamic model: assessment and prospect. Biol Rev Camb Philos Soc 92(2), 830-853. DOI: 10.1111/brv.12256
Triantis, K. A., Vardinoyannis, K., Tsolaki, E. P., Botsaris, I., Lika, K., Mylonas, M. (2006) Re‐approaching the small island effect. Journal of Biogeography, 33(5), pp.914–923.
van der Geer, Alexandra A. E., Lomolino, Mark V. & Lyras, George A. (2017) ‘Island Life’ before man: biogeography of palaeo‐insular mammals. Journal Of Biogeography, 44(5), pp.995–1006.
Vitousek, P.M. (2002) Oceanic islands as model systems for ecological studies. Journal of Biogeography, 29(5‐6), pp.573–582.
Whittaker, Robert J., Triantis, Kostas A. & Ladle, Richard J. (2008) general dynamic theory of oceanic island biogeography. Journal of biogeography, 35(6), pp.977–994.
Whittaker, R.J., Fernández-Palacios, J.M., Matthews, T.J., Borregaard, M.K. and Triantis, K.A., 2017. Island biogeography: Taking the long view of nature’s laboratories. Science, 357(6354)
#Write for Routes
Are you 6th form or undergraduate geographer?
Do you have work that you are proud of and want to share?
Submit your work to our expert team of peer reviewers who will help you take it to the next level.
Related articles
